Article Text

Abstract
Objectives Recent studies have identified vaginal bacterial taxa associated with increased HIV risk. A possible mechanism to explain these results is that individual taxa differentially promote cervicovaginal inflammation. This study aimed to explore relationships between concentrations of bacteria previously linked to HIV acquisition and vaginal concentrations of proinflammatory cytokines and chemokines.
Methods In this cross-sectional analysis, concentrations of 17 bacterial taxa and four proinflammatory cytokines (interleukin (IL)-1β, IL-6, IL-10 and tumour necrosis factor alpha (TNFα)) and two proinflammatory chemokines (IL-8 and interferon gamma-induced protein 10) were measured in vaginal swabs collected from 80 HIV-uninfected women. Cytokine and chemokine concentrations were compared between women with bacterial concentrations above or below the lower limit of detection as determined by quantitative PCR for each taxon. Principal component analysis was used to create a summary score for closely correlated bacteria, and linear regression analysis was used to evaluate associations between this score and increasing concentrations of TNFα and IL-1β.
Results Detection of Dialister micraerophilus (p=0.01), Eggerthella sp type 1 (p=0.05) or Mycoplasma hominis (p=0.03) was associated with higher TNFα concentrations, and detection of D. micraerophilus (p<0.01), Eggerthella sp type 1 (p=0.04), M. hominis (p=0.02) or Parvimonas sp type 2 (p=0.05) was associated with significantly higher IL-1β concentrations. Seven bacterial taxa (D. micraerophilus, Eggerthella sp type 1, Gemella asaccharolytica, Sneathia sp, Megasphaera sp, M. hominis and Parvimonas sp type 2) were found to be highly correlated by principal component analysis (eigenvalue 5.24, explaining 74.92% of variability). Linear regression analysis demonstrated associations between this principal component and concentrations of TNFα (β=0.55, 95% CI 0.01 to 1.08; p=0.048) and IL-1β (β=0.96, 95% CI 0.19 to 1.74; p=0.016).
Conclusions This study provides evidence that several highly correlated vaginal bacterial taxa may influence vaginal cytokine and chemokine concentrations. These results suggest a mechanism where the presence of specific bacterial taxa could influence HIV susceptibility by increasing vaginal inflammation.
- vaginal microbiology
- sexual health
- Trichomonas
- immunology
- HIV women
Statistics from Altmetric.com
Introduction
The vaginal microbiota likely exists on a continuum between ‘optimal’ states, characterised by Lactobacillus sp predominance, and ‘suboptimal’ states, distinguished by increased anaerobic bacteria, species richness and species diversity.1–3 Bacterial vaginosis (BV) is a clinical syndrome distinguished by increased detection of suboptimal bacterial taxa, and has been associated with elevated risk of HIV acquisition.2 4 5 One possible explanation for this association is that BV causes a proinflammatory state, resulting in recruitment of activated immune cells and breakdown in the mucosal barriers of the cervix and vagina.5–7 Assessment of vaginal inflammation in women has consistently demonstrated an association between BV and interleukin (IL)-1β, but associations between BV and other proinflammatory cytokines and chemokines have varied between studies.1
BV does not represent a single microbiological entity, and the bacterial communities in BV are heterogeneous.8 One plausible explanation for the variation in cytokine/chemokine patterns noted in prior reports is that cytokines and chemokines are influenced by individual bacterial taxa, which may differ between women with BV.8 For example, a recent publication demonstrated that Gardnerella vaginalis and Atopobium vaginae were associated with decreased concentrations of interferon gamma-induced protein 10 (IP-10) and increased concentrations of IL-8, IL-1α, IL-1β and IL-12.9 A growing body of evidence supports the role of individual taxa in influencing clinical outcomes, such as HIV acquisition.10 Recently, our group demonstrated an association between risk of HIV-1 acquisition in women and concentrations of seven bacterial taxa associated with suboptimal vaginal states, including Eggerthella sp type 1, Gemella asaccharolytica, Sneathia sp, Megasphaera sp, Mycoplasma hominis, Parvimonas sp type 1 and Parvimonas sp type 2.10 To better understand which bacterial taxa are associated with inflammation, concentrations of proinflammatory cytokines, chemokines and select bacterial taxa were measured in vaginal fluid. In addition, principal component analysis (PCA) and linear regression were performed to assess patterns in bacterial concentrations and associations between those patterns and cytokine and chemokine levels. Together, these data provide insight into the potential mechanisms by which bacterial taxa may influence cervical inflammation and HIV risk.
Materials and methods
Study design and participants
A secondary cross-sectional analysis of data collected from the Mombasa Cohort11 and the Mama Salama Study 12 was performed. Detailed information on both cohorts and study procedures has been described.11 12 In brief, the Mombasa Cohort is a longitudinal, open cohort study of female sex workers in Mombasa, Kenya, and the Mama Salama Study was a prospective study of HIV-negative, pregnant women presenting to the Ahero Sub-District Hospital or Bondo District Hospital in Kenya.11 12 Country-specific and investigator-affiliated ethical review board approval was obtained for both studies. All participants provided written informed consent.
After enrolment, women in both cohorts returned for follow-up visits every 1–3 months for collection of behavioural and demographic data, physical examination and testing for STIs, as previously described.10–12 At specific follow-up visits defined for each cohort, vaginal swabs were collected for quantitative PCR (qPCR), analysis of bacteria and measurement of cytokines and chemokines. This cross-sectional analysis includes data from a single visit per woman at which both microbiota and cytokine/chemokine data were available. Among women who became infected with HIV, the visit selected was the last visit prior to HIV seroconversion, as previously described.10 One woman who was HIV positive and one woman with gonococcal cervicitis at the time of sampling were excluded, as these infections may alter the relationships between microbiota and cytokines.13 14 Data on herpes simplex virus (HSV) serostatus or HSV shedding were not available for the majority of women and are not included in this analysis. Due to differences in the timing of vaginal swab collection and STI testing per study protocols, assessment for Chlamydia trachomatis (n=19) and genital ulcer disease (n=50) was only performed on a subset of women at the analysis visit.
Laboratory procedures
STItesting
Testing for HIV was performed by ELISA in the Mombasa Cohort using the Pishtaz HIV 1.2 ELISA (Pishtaz Teb Diagnostics, Tehran, Iran) for HIV screening and the Vironostika HIV-1 Uni-Form II Ag/Ab (bioMérieux, Marcy I’Etoile, France) for confirmatory testing.10 11 In the Mama Salama S tudy, the first-generation Gen-Probe HIV viral load assay (Hologic/Gen-Probe, San Diego, CA, USA) was used for HIV testing.10 12 The Gen-Probe APTIMA Combo-2 Assay (Hologic/Gen-Probe) was used in both cohorts for diagnosis of infection with Neisseria gonorrhoeae and C. trachomatis.11 12 For both cohorts, BV was diagnosed by Gram stain according to the method of Nugent and Hillier, and Trichomonas vaginalis infection was diagnosed by wet preparation.11 12 15
Vaginal sample collection
Vaginal samples were collected during speculum-assisted pelvic examination (Mombasa Cohort) or by self-collection (Mama Salama Study) using push-off Dacron swabs from FitzCo, Inc (Spring Park, MN, USA). Vaginal swabs were stored at −80°C in Kenya, shipped on dry ice to Seattle, then stored at −80°C at the Fred Hutchison Cancer Research Center in Seattle, WA, until use.
Quantitative PCR
DNA extraction and bacterium-specific qPCR from vaginal fluid samples were performed for the following bacterial taxa according to published protocols: Aerococcus christensenii, A. vaginae, BV-associated bacterium 2 (BVAB2), Dialister micraerophilus, Dialister sp type 2, Eggerthella sp type 1, G. vaginalis, G. asaccharolytica, Lactobacillus crispatus, Sneathia sp, Megasphaera sp, M. hominis, Parvimonas sp type 1, Parvimonas sp type 2, Porphyromonas asaccharolytica/uenonis, Porphyromonas sp type 1, Porphyromonas bennonis and Prevotella genus.8 10 16 17
Measurement of vaginal cytokines and chemokines
Levels of IL-1β, IL-6, IL-8, IL-10, tumour necrosis factor alpha (TNFα) and IP-10 in vaginal samples were assessed using a V-Plex Custom Human Cytokine panel from Meso Scale Discovery (Rockville, MD, USA) following the manufacturer’s instructions. Cytokine and chemokine values for samples with levels below the lower limit of detection (LLD) were set to the midpoint between zero and the LLD for that cytokine.
Statistical analysis
For this cross-sectional analysis, data from the visit at which vaginal samples were collected were used to define the population characteristics. Demographic and behavioural data were reported using descriptive statistics. The primary exposure was dichotomised as detection of bacterial taxa above or below the LLD of qPCR. Bacterial taxa analysed included: (1) group 1: seven bacterial taxa recently reported to have significant concentration-dependent associations with increased risk of HIV acquisition (Eggerthella sp type 1, G. asaccharolytica, Sneathia sp, Megasphaera sp, M. hominis, Parvimonas sp type 1 and Parvimonas sp type 2)10; (2) group 2: ten additional taxa that demonstrated a statistical trend towards association with HIV acquisition by rank abundance in a prior analysis, including: A. christensenii, A. vaginae, BVAB2, D. micraerophilus, Dialister sp type 2, G. vaginalis, P. asaccharolytica/uenonis, Porphyromonas sp type 1, P. bennonis and Prevotella genus10; and (3) L. crispatus, a well-described marker of vaginal health.1–3 Secondary analyses were conducted using the log10 concentration of bacterial taxa as the exposure. The primary outcome was the log2-transformed concentration of IL-1β, IL-6, IL-8, IL-10, TNFα and IP-10. Transformation to the log2 scale was performed to normalise cytokine/chemokine concentrations and increase biological relevance, so that a one-unit change corresponds to a doubling (1 log2 increase) or halving (1 log2 decrease) of concentration.
For each taxon, cytokine/chemokine concentrations in participant samples with and without bacterial detection were compared using Wilcoxon rank-sum tests. All group 1 bacterial taxa were carried forward for further analysis a priori. Each of the group 2 bacterial taxa were carried forward only if associated with the cytokines or chemokines of interest at p<0.10. Of this set of exposure variables, only one group 2 bacterial taxon, D. micraerophilus, met this criterion and was carried forward for PCA.
Due to significant correlations between the different bacterial taxa studied, and to reduce the dimensionality of the data set, PCA was performed on log10-transformed concentrations of the bacterial taxa carried forward as described above. One factor had an eigenvalue of 5.39 and accounted for 67.4% of variability in the analysis; no other factors had eigenvalues >1. Parvimonas sp type 1 concentration had a uniqueness score of 82.62%, suggesting that this taxon did not share similar features with the other bacterial taxa in the model, and was subsequently removed. The seven remaining bacterial taxa (D. micraerophilus, Eggerthella sp type 1, G. asaccharolytica, Sneathia sp, Megasphaera sp, M. hominis and Parvimonas sp type 2) underwent repeat PCA, confirming generation of a single factor of highly correlated bacterial species (eigenvalue 5.24, explaining 74.92% of variability). The high degree of variability explained by this factor suggests that these taxa are highly correlated, and are better analysed as a single variable representing the degree of suboptimal taxa present rather than as individual predictors assumed to be independent. Therefore, results of the PCA were used to generate a principal component score (labelled ‘suboptimal taxa score’) for each participant. This score is interpretable as a summary statistic generated for each woman based on the relative concentrations of the seven bacterial taxa included in the PCA.
Linear regression was performed to determine if the suboptimal taxa score and other components of the vaginal microbiota were independently associated with log2-tranformed concentrations of TNFα and IL-1β. Primary predictors included the suboptimal taxa score and Parvimonas sp type 1 concentration. Potential confounders included hormonal status (categorised as pregnant, no hormonal contraception or use of hormonal contraception), vaginal washing (defined as washing beyond the introitus in the week prior to the analysis visit), age, number of sex partners, frequency of unprotected sex, frequency of vaginal sex, T. vaginalis infection, genital ulcer disease and pregnancy status (pregnant vs not pregnant). Hormonal status and vaginal washing were selected a priori for inclusion in multivariate analysis based on review of the literature suggesting that both may influence vaginal inflammation.18 19 The remaining potential confounders were evaluated by linear regression for associations with TNFα or IL-1β, and included in multivariable modelling if associated with the cytokine of interest at p<0.10.
In addition to the above analyses, the relationship between diagnosis of BV (Nugent score ≥7) or abnormal microbiota (Nugent score ≥4) and cytokine, chemokine or bacterial taxa concentration was evaluated using Wilcoxon rank-sum tests. Pearson’s correlation test was also performed to assess for correlations between the suboptimal taxa score, Nugent score, TNFα and IL-1β. Statistical analyses were conducted using Stata V.15.1 (College Station, Texas, USA).
Results
Paired vaginal bacterial qPCR and cytokine data were available for 80 eligible women, 50 from the Mama Salama Study, of whom 19 (23.75%) were pregnant, and 30 from the Mombasa Cohort. Median age at the time of sample collection used in this analysis was 24.5 years (IQR 19–35). Three women tested positive for T. vaginalis (3.8%) and 27 of 78 women (34.6%) for whom Gram stain results were available were diagnosed with BV (Nugent score ≥7). Additional demographic and health characteristics are reported in table 1.
Characteristics at the time of sample collection for 80 participating women
Figure 1 shows the relationship between log2 cytokine or chemokine concentrations and detection of bacterial taxa selected for analysis. Detection of D. micraerophilus (p=0.01), Eggerthella sp type 1 (p=0.05) or M. hominis (p=0.03) was associated with higher concentrations of TNFα. Similarly, detection of D. micraerophilus (p<0.01), Eggerthella sp type 1 (p=0.04), M. hominis (p=0.02) or Parvimonas sp type 2 (p=0.05) was associated with higher concentrations of IL-1β. In contrast, detection of L. crispatus was associated with lower concentrations of TNFα (p=0.04) and IL-1β (p=0.04). Detection of Megasphaera sp was associated with decreased concentrations of IP-10 (p=0.027). Because multiple bacterial taxa were associated with TNFα and IL-1β, and to restrict the total number of statistical comparisons, subsequent analysis focused on these two cytokines.

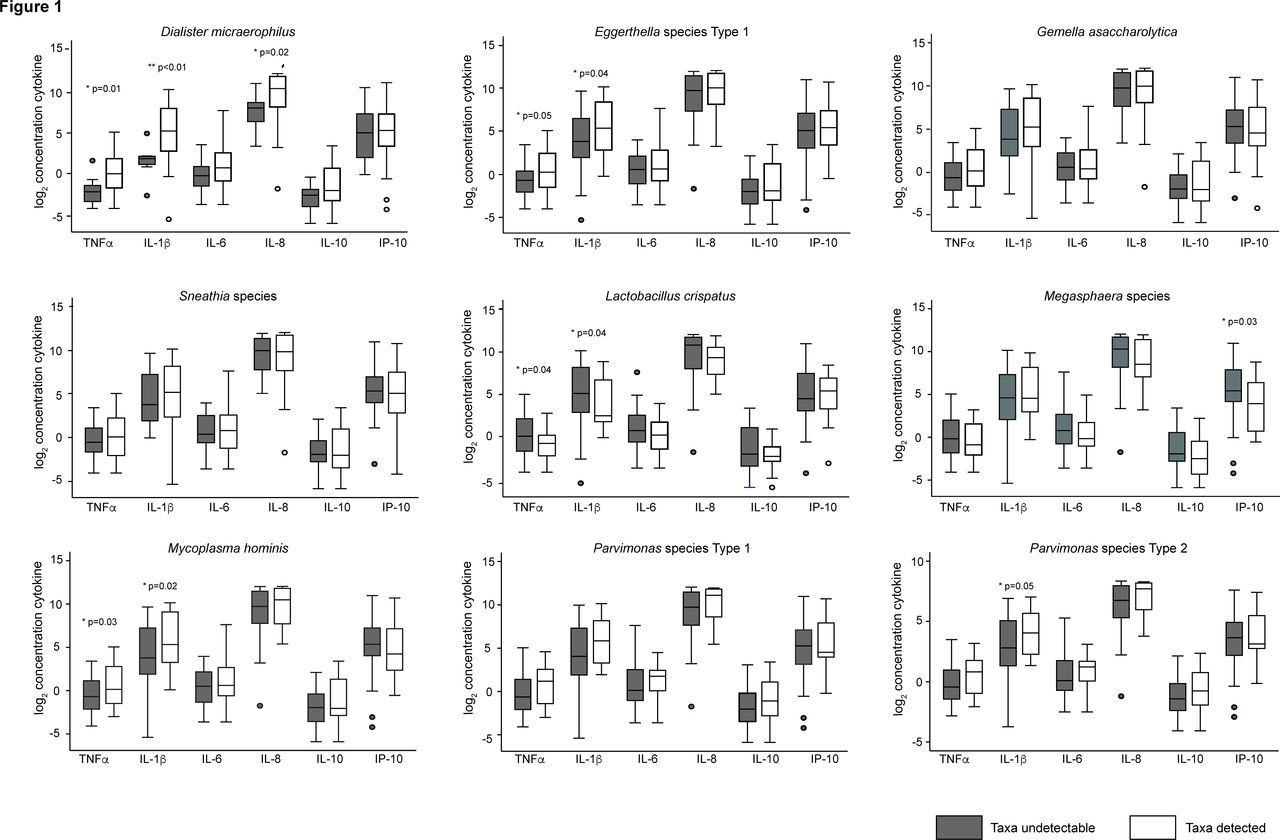{kind=link}
Box plots of log2-transformed cytokine level by detection of bacterial taxa. The log2 of cytokine concentrations when bacterial taxa were below (dark bars) or above (white bars) the lower limit of detection. P values were calculated using the Wilcoxon rank-sum test. The lower limits of detection for each cytokine are as follows: IL-1β, 0.048 pcg/mL; IL-6, 0.164 pcg/mL; IL-8, 0.090 pcg/mL; IL-10, 0.033 pcg/mL; TNFα, 0.118 pcg/mL; IP-10, 0.109 pcg/mL. IL, interleukin; IP-10, interferon gamma-induced protein 10; TNFα, tumour necrosis factor alpha.
Table 2 presents the results of bivariable and multivariable linear regression using log2-transformed TNFα concentration as the outcome. Higher values of the suboptimal taxa score (β=0.56, 95% CI 0.06 to 1.07; p=0.029) were associated with higher TNFα concentrations. In contrast, higher concentrations of Parvimonas sp type 1 were not associated with higher TNFα concentrations (β=0.34, 95% CI −0.12 to 0.79; p=0.14). The association of the suboptimal taxa score (β=0.55, 95% CI 0.01 to 1.08; p=0.048) with higher TNFα concentrations remained statistically significant in analyses adjusted for T. vaginalis, hormonal status, vaginal washing and age.
Cofactors for log2 TNFα concentration
Table 3 presents the results of bivariable and multivariable linear regression using log2-transformed IL-1β concentration as the outcome. Unadjusted analysis demonstrated that higher values of the suboptimal taxa score (β=0.97, 95% CI 0.25 to 1.70; p=0.009) were associated with higher concentrations of IL-1β. Higher concentrations of Parvimonas sp type 1 were not associated with higher concentrations of IL-1β (β=0.43, 95% CI −0.23 to 1.09; p=0.20). In multivariable analysis adjusting for T. vaginalis, hormonal status, vaginal washing and age, the suboptimal taxa score remained significantly associated with IL-1β concentration (β=0.96, 95% CI 0.19 to 1.74; p=0.016).
Cofactors for log2 IL-1β concentration
Pearson’s correlation test demonstrated statistically significant correlations between the suboptimal taxa score and Nugent score (p<0.001), TNFα (p=0.030) and IL-1β (p=0.009) (online supplementary figure 1). No significant associations were found between diagnosis of BV and concentrations of any cytokines or chemokines tested, including TNFα and IL-1β (online supplementary table 1); increasing concentrations of IL-1b were associated with the presence of abnormal microbiota (Nugent score ≥4) (online supplementary table 2). However, diagnosis of BV was associated with the suboptimal taxa score (p<0.001). Diagnosis of BV was also associated with higher concentrations of all bacterial taxa associated with HIV acquisition except M. hominis, and with lower concentrations of L. crispatus (online supplementary table 1). Sensitivity analysis excluding the three women with T. vaginalis infections did not change the overall results (data not shown).
Supplemental material
Supplemental material
Supplemental material
Discussion
In this exploratory cross-sectional analysis, detection of D. micraerophilus, Eggerthella sp type 1 or M. hominis was associated with higher concentrations of TNFα, while detection of G. asaccharolytica, Eggerthella sp type 1, M. hominis or Parvimonas sp type 2 was associated with higher concentrations of IL-1β. PCA highlighted an association between a principal component reflecting correlated suboptimal bacterial taxa and higher concentrations of TNFα and IL-1β.
TNFα is a proinflammatory cytokine with multiple functions, including activation of neutrophils and macrophages.20 Studies performed using in vitro coculture models have consistently shown higher concentrations of TNFα in the presence of bacteria associated with BV.4 6 On the other hand, results of clinical studies examining the association between BV and TNFα have been mixed.4 6 21–23 In this analysis, some bacterial taxa were associated with higher concentrations of TNFα, while others were not. Interestingly, the principal component score of highly correlated bacterial taxa, several of which were individually associated with TNFα, was associated with higher TNFα concentrations, while a diagnosis of BV was not. Together, these data suggest that vaginal TNFα concentration may vary based on the specific composition of the vaginal microbiota rather than the presence or absence of clinical BV.
IL-1β is produced as an inactive precursor by multiple cell types and functions to activate CD4+ T cells and generate proinflammatory cytokines.24 Numerous studies have shown an association between BV and IL-1β, and treatment of BV decreases levels of IL-1β.1 25 This analysis demonstrates that detection of several individual bacterial taxa (D. micraerophilus, Eggerthella sp type 1, M. hominis or Parvimonas sp type 2) is associated with higher IL-1β concentrations, and that a principal component that comprised highly correlated bacterial taxa was also associated with higher IL-1β concentrations after adjustment for potential confounders. As with TNFα, diagnosis of BV was not associated with IL-1β, suggesting that IL-1β expression may be highly dependent on the presence of specific bacteria, either alone or in combination.
The suboptimal vaginal bacterial taxa analysed here have been associated with increased risk of HIV acquisition.10 One hypothesis to explain this increased risk is that these bacteria recruit CD4+ T cells to the site of HIV entry by inducing a proinflammatory state.26 In this study, these high-risk bacterial taxa were associated with elevated levels of TNFα and IL-1β, which may promote HIV acquisition via a number of pathways. For example, in vitro studies suggest that TNFα signalling disrupts vaginal mucin production, which may facilitate HIV entry by decreasing epithelial integrity.6 27 Similarly, exposure to bacteria that increase levels of both cytokines has been associated with upregulation of nuclear factor-kappa B and other proinflammatory pathways in vaginal tissue models.6 However, given the difficulty of culturing some vaginal bacteria, there is an incomplete understanding of the bacterial antigens that trigger this signalling cascade. It is also possible that other mechanisms are involved. For example, individual bacterial taxa may secrete a factor that promotes HIV replication,26 or alter the vaginal mucosal barrier via other mechanisms, such as direct cytoskeletal disruption or alteration of proteolytic activity at the epithelial surface.28
This study had a number of strengths. The use of taxon-specific qPCR allowed for examination of individual bacterial taxa concentrations and vaginal cytokine concentrations, which is becoming increasingly important as more studies report associations between individual bacterial taxa and adverse outcomes.5 10 Generation of a principal component facilitated analysis of bacterial taxa that were shown to be highly correlated, and provided information about how highly correlated taxa influence cytokine production in concert. These results should also be interpreted in the setting of several limitations. As a secondary analysis, the data presented should be considered hypothesis generating, and a prospective study to address changes in cytokines before and after treatment of BV will be necessary to prove causality. Future studies of the relationship between individual bacterial taxa and chemokines involved in T cell recruitment (ie, CCL5, MIP-1α) will also be critical for developing a mechanistic understanding of the role of specific taxa in HIV acquisition.29 Furthermore, this study may not have had the power to detect small differences in cytokine concentrations. Additionally, data were not available for HSV-2, which is associated with both inflammation and HIV acquisition, and could act as a confounder of the association between vaginal bacteria and inflammation.30 31 Similarly, only 19 women were tested for C. trachomatis at the visits included in this study. However, women in the Mombasa Cohort were tested multiple times during their participation, and C. trachomatis was treated when detected. Women in the Mama Salama Study were only tested and treated at the baseline visit for C. trachomatis, however the overall prevalence was only 6%, suggesting this is a relatively infrequent diagnosis in this population.12 Therefore, the number of untreated cases of chlamydia should have been low.
In summary, this analysis demonstrates associations between individual bacterial taxa and proinflammatory cytokines, suggesting that individual bacterial taxa may play an important role in determining the inflammatory state of the vagina. Future studies focusing on changes in inflammation if these bacteria are eliminated could help strengthen the evidence for a causal relationship between vaginal bacteria and inflammation, as well as demonstrate the potential of vaginal health approaches for reducing HIV susceptibility.
Key messages
Several bacterial taxa associated with HIV acquisition were associated with higher concentrations of tumour necrosis factor alpha (TNFα) and interleukin (IL)-1β.
A score derived from principal component analysis of these highly correlated bacterial taxa was associated with higher Nugent score and with higher concentrations of TNFα and IL-1β.
Bacterially mediated inflammation in the vagina is one mechanism that could account for increased HIV risk in women.
Acknowledgments
We would like to thank all of the women who participated in the Mama Salama Study and Mombasa Cohort. We would also like to acknowledge the clinical, laboratory and administrative staff at each study site for their dedication and assistance with this project.
References
Footnotes
Handling editor Jo-Ann Passmore
Contributors ALD, JK, DM, BAR, JO, RSM and GJS were involved in design of the parent studies, and SMG conceived the present analysis. JK, ALD, DM, LO and GJS performed data collection. MCS, DAL, BW, BAR, JO and SMG participated in data analysis. DNF and SS were involved in design and implementation of quantitative PCR procedures. TLF and MMM performed qPCR data collection and analysis. BW and DAL generated cytokine data. JO, GJS, RSM and DNF acquired funding for this study. BAR provided statistical guidance. MCS and SMG wrote the initial draft of the manuscript. All authors contributed to the final draft of the manuscript and approved the final draft for submission.
Funding This study was supported by the National Institute of Child Health and Human Development of the National Institutes of Health (NIH P01-HD64915). Data and sample collection in the Mombasa Cohort were supported through the National Institute of Allergy and Infectious Diseases of the NIH (NIH R37 AI38518). The Mombasa research site receives infrastructure support from the University of Washington Center for AIDS Research (NIH P30-AI27757). RSM receives funding for mentoring through NIH K24 HD88229. ALD receives support from K01 AI116298. MCS is supported by the T32 Host Defense Training grant (NIH 5T32AI007044-43 PI, van Voorhis) as an infectious disease fellow. SMG was supported by the Robert W Anderson Endowed Professorship in Medicine.
Competing interests RSM receives research funding, paid to the University of Washington, from Hologic. TLF has a patent, Molecular Diagnosis of Bacterial Vaginosis, licensed to Becton Dickinson. SS, MMM, DAL and TLF report grants from the NIH during the conduct of the study. All other authors have nothing to disclose.
Patient consent for publication Not required.
Ethics approval Kenyatta National Hospital (KNH) Ethics Review Committee (ERC), application number: P197/04/2012–University of Washington (UW) Human Subjects Committee, IRB number STUDY00000615-KNH ERC, application number: P114/04/2010, renewed in 2018–UW Human Subjects Committee, IRB number 38472.
Provenance and peer review Not commissioned; externally peer reviewed.
Data sharing statement All data relevant to the study are included in the article or uploaded as supplementary information.