Article Text

Abstract
Objectives Complex interactions of vaginal microorganisms with the genital tract epithelium shape mucosal innate immunity, which holds the key to sexual and reproductive health. Bacterial vaginosis (BV), a microbiome-disturbance syndrome prevalent in reproductive-age women, occurs commonly in concert with trichomoniasis, and both are associated with increased risk of adverse reproductive outcomes and viral infections, largely attributable to inflammation. To investigate the causative relationships among inflammation, BV and trichomoniasis, we established a model of human cervicovaginal epithelial cells colonised by vaginal Lactobacillus isolates, dominant in healthy women, and common BV species (Atopobium vaginae, Gardnerella vaginalis and Prevotella bivia).
Methods Colonised epithelia were infected with Trichomonas vaginalis (TV) or exposed to purified TV virulence factors (membrane lipophosphoglycan (LPG), its ceramide-phosphoinositol-glycan core (CPI-GC) or the endosymbiont Trichomonas vaginalis virus (TVV)), followed by assessment of bacterial colony-forming units, the mucosal anti-inflammatory microbicide secretory leucocyte protease inhibitor (SLPI), and chemokines that drive pro-inflammatory, antigen-presenting and T cells.
Results TV reduced colonisation by Lactobacillus but not by BV species, which were found inside epithelial cells. TV increased interleukin (IL)-8 and suppressed SLPI, likely via LPG/CPI-GC, and upregulated IL-8 and RANTES, likely via TVV as suggested by use of purified pathogenic determinants. BV species A vaginae and G vaginalis induced IL-8 and RANTES, and also amplified the pro-inflammatory responses to both LPG/CPI-GC and TVV, whereas P bivia suppressed the TV/TVV-induced chemokines.
Conclusions These molecular host–parasite–endosymbiont–bacteria interactions explain epidemiological associations and suggest a revised paradigm for restoring vaginal immunity and preventing BV/TV-attributable inflammatory sequelae in women.
- TRICHOMONAS
- VAGINAL MICROBIOLOGY
- IMMUNOLOGY
- BACTERIAL VAGINOSIS
- WOMEN
This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 3.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/3.0/
Statistics from Altmetric.com
Introduction
Homeostasis of the genital tract mucosa in reproductive-age women is challenged by resident microorganisms that vary depending on menstrual cycle, contraceptive and vaginal practices, and sexually transmitted pathogens.1–3 Culture techniques as well as modern genetic and metagenomic methods support the paradigm that both type and relative abundance of different bacterial taxa are indicative of vaginal health.4–7 A deviation from the norm, including increased numbers of certain species (eg, Gardnerella vaginalis, Atopobium vaginae and Prevotella bivia) and decreased abundance of Lactobacillus spp., has been coined as the syndrome of disturbed vaginal microbiota or bacterial vaginosis (BV).4–6 ,8 BV is among the most common conditions seen by primary care physicians in reproductive-age women9 and is often combined with infection by the flagellated protozoan Trichomonas vaginalis (TV).10 The occurrence rates and prevalence of BV and TV are believed to be underestimated because both are commonly asymptomatic and neither requires mandatorily reporting.
Common features of BV and TV are lack of effective adaptive immunity, high recurrence rate, high risk of reproductive problems (eg, chorioamnionitis, preterm birth, premature rupture of membranes, low birth weight, pelvic inflammatory disease) and failure of antibiotic treatment to break the inflammatory sequelae that complicate maternal–fetal interactions.11 Another common adverse outcome of both BV and TV is increased risk of HIV and other sexually transmitted infections.11 Both BV and TV are associated with vaginal discharge and abnormal levels of cytokines in cervicovaginal secretions.11 Although epidemiological findings link trichomoniasis to low abundance of lactobacillus, causative relationships between TV and BV, and their combined impact on vaginal immunity, have not been elucidated.10
We hypothesised that the TV parasite selectively interferes with patterns of vaginal colonisation and that vaginal bacteria reciprocally modify the mucosal immune balance and responses to TV virulence determinants, including its predominant surface lipophosphoglycan (LPG),12 ,13 also named lipoglycan,14 its ceramide-phosphoinositol-glycan core (CPI-GC), responsible for parasite adherence and chemokine response,12 ,13 and the endosymbiotic dsRNA viruses carried by TV (Trichomonas vaginalis virus (TVV)) that are sensed by the human host, causing inflammatory responses, magnified by antibiotic treatment.15 Here we newly applied a bacterial colonisation model16 ,17 to test this hypothesis.
Methods
Epithelial cells
Epithelial cell lines Vk2/E6E7, Ect1/E6E7 and End1/E6E7, representing normal human vagina, ectocervix and endocervix,18 were maintained in keratinocyte serum-free medium (KSFM) supplemented with 50 μg/mL bovine pituitary extract, 0.1 ng/mL epidermal growth factor, penicillin/streptomycin (all from Invitrogen) and CaCl2 (Fisher).18 Antibiotics were omitted in all experiments. Ect1/E6E7 and End1/E6E7 were derived from the same woman thus allowing an isogenic comparison of simple versus stratified non-keratinising cell types.18 All three cell lines, previously compared with their progenitors and a non-transformed organotypic ectocervical-vaginal tissue model, have shown immune responses to bacteria and a variety of pathogenic determinants including toll-like receptor (TLR) ligands, LPG and TVV, similar to those by their primary counterparts.12 ,15 ,17–22
TV LPG and CPI-GC
UR1, a TV isolate that carries three TVV species, was previously obtained from a symptomatic patient, cloned and cultured in Diamond's modified media as described.15 We chose this isolate for our experiments because it represents the majority (>80%) of those in our collection in being infected with multiple TVV species.15 For LPG extraction, parasites were harvested in late log phase (24 h) by centrifugation, washed twice with phosphate-buffered saline (PBS, pH 7.4) (Invitrogen), consecutively extracted with methanol/chloroform (1:2) and solvent E and purified on an octyl-Sepharose column.13 The CPI-GC core was released by mild acid treatment (100 mM TFA) of LPG as described.13 LPG and CPI-GC purity and lack of endotoxin contamination were confirmed as described.13 Both LPG and CPI-GC were used at a dose of 240 μg/ml based on previously established lack of toxicity, peak of inflammatory dose response and corresponding relevant multiplicity of infection.13
TVV virions
TVV1 virions were purified, identified by gel electrophoresis and assessed for intact structure by electron microscopy after negative staining with uranyl formate as described in detail elsewhere.15 Virion concentrations were estimated by BioRad protein assay against a bovine serum albumin standard and assuming 120 capsid protein molecules (75 kDa) per virion. Virions were stored in aliquots at −80°C until used in stimulation experiments at ∼1×1011 virions/cm2 of epithelial surface.15
Bacterial colonisation and TV co-infection
Lactobacillus acidophilus, Lactobacillus crispatus, Lactobacillus jensenii, G vaginalis and P bivia were originally isolated by vaginal swabs.23 These isolates were identified using established phenotypic criteria and identification was confirmed using the Microbial Identification System for long chain fatty acid analysis (MIDI Inc).23 For this study, we chose Lactobacillus spp. that are characteristically common and/or stabilise the vaginal microbiota in women without BV, for example, L crispatus and L jensenii.24 ,25 The L acidophilus strain was chosen because it has shown non-inflammatory homeostatic properties and represents one of the few taxa that have thus far shown promise in randomised trials for the cure of BV.26 A vaginae was acquired from the American Type Culture Collection (BAA-55). Bacterial suspension in antibiotic-free KSFM medium were added to confluent epithelial monolayers (at 2.2×106 CFU/cm2 for all bacteria except G vaginalis which was highly virulent and used at a 10-fold lower dose) for 24 h under anaerobic conditions to allow for colonisation.17 After 24 h, non-adherent or loosely attached bacteria were removed with the culture medium. TV (1.25×105 parasites/cm2), LPG (240 µg/ml) or CPI-GC (240 µg/ml) were then added, and epithelia-associated bacteria as well as immune responses were assessed after another 24 h. Synthetic TLR2/TLR6 ligand MALP-2 (25 nM; Alexis Biologicals) analogue of Mycoplasma fermentans lipopeptide macrophage activating lipopeptide was used as a pro-inflammatory control. At the end of each stimulation period, supernatants were unless specified collected for soluble mediators and epithelial cells were lysed for colony-forming units (CFU) counts, used for viability assessment or used for microscopy.16 ,17 For CFU counts, epithelial cells were washed twice with PBS and hypotonically lysed in HyPure water (Fisher) for 15 min. The lysates were seeded on agar following adjustment of osmolarity by adding equal volume of 2× PBS as described.17 Bacterial suspensions were also allowed to grow in parallel in the absence of epithelial cells in the presence or absence of TV to determine the initial multiplicity of infection for each microorganism, the degree of growth of each during the experiment and direct effects of TV. In some experiments, epithelia-free bacterial suspensions or colonised epithelial monolayers were exposed to a combination of 100 U/mL penicillin and 100 µg/mL streptomycin (both from Invitrogen) or 100 µM metronidazole (Acros Organics) previously shown to be non-toxic to epithelial cells15 and incubated for 4 h followed by plating on agar for relative assessment of extracellular and intracellular CFU.
Cell viability
Epithelial cell viability in the presence of LPG, CPI-GC, MALP-2 or TVV virions was assessed by the non-radioactive CellTiter96 MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide) assay (Fisher). Cell viability in the presence of TV and bacteria was assessed microscopically and by the trypan blue exclusion assay (Fisher) because the microorganisms also convert the MTT dye and thus the MTT assay is not an optimal choice for coculture conditions.
Inflammation-associated proteins
The following chemokines were measured in culture supernatants using electrochemiluminescence multiplex assays on a Sector Imager 2400 (Meso Scale Discovery): interleukin-8 (IL-8, CXCL8), chemoattractant primarily for neutrophils; regulated and normal T cell expressed and secreted protein (RANTES, CCL5), chemoattractant primarily for T cells; macrophage inflammatory protein-3α (MIP-3α, CCL20), chemoattractant primarily for dendritic cells; and interferon γ-induced protein 10 (IP-10, CXCL10), chemoattractant for T cells, monocytes, NK cells and dendritic cells. Secretory leucocyte protease inhibitor (SLPI) levels were measured by Quantikine ELISA (R&D Systems).
Statistics
Data were analysed by analysis of variance (ANOVA) (GraphPad Prism, V.5.0). p Values <0.05 were considered significant.
Results
Inoculation of live TV reduced the CFU numbers of epithelia-associated L acidophilus and L jensenii by more than two log10 values (figure 1A). L crispatus and G vaginalis showed a trend of decrease, which however did not reach a log10 difference. Live TV had no effect on P bivia or A vaginae. LPG, CPI-GC and TVV had no significant effects except that epithelia-associated G vaginalis was slightly increased by CPI-GC. These data suggested that the live TV was responsible for the observed reduction in epithelia-associated bacterial CFU numbers.

Effect of Trichomonas vaginalis (TV) on bacterial colonisation in an in vitro cervicovaginal colonisation model. (A) Colony-forming units (CFU) associated with endocervical epithelial cells 48 h postcolonisation and 24 h after exposure to live TV or purified TV virulence factors. Bars represent means±SEM of duplicate measurements representing consistent results from five independent experiments. ***p<0.001, **p<0.01, *p<0.05, live TV, lipophosphoglycan, ceramide-phosphoinositol-glycan core and Trichomonas vaginalis virus virions different from medium control, two-way ANOVA, Bonferroni post-test. Similar results were obtained with vaginal and ectocervical cells (data not shown). (B) Electron micrographs from the vaginal colonisation model. Arrows in the higher magnification image (upper panel) indicate adherent or internalised Atopobium vaginae in vaginal epithelial cells. (C–H) CFU numbers assessed after 4 h antibiotic treatment of epithelia-free bacterial suspensions (C–E) or vaginal epithelial monolayers (F–H) precolonised with bacteria for 24 h. Data are means±SEM of duplicate cultures representing one of three experiments. med, antibiotic-free medium, p/s, combined penicillin–streptomycin; metro, metronidazole, Vk, vaginal epithelial cells. *p<0.05, **p<0.01, ***p<0.001, antibiotics different from no antibiotics, +p<0.05, ++p<0.01, +++p<0.001, supernatant different from lysate. (I) Reduction of planktonic or epithelia-associated CFU after 24 h infection with TV. Data are means and SEM from duplicate cultures of vaginal (Vk), ectocervical (Ect) and endocervical (End) epithelial cells representing two independent experiments. *p<0.05, **p<0.01, ***p<0.001, TV—infected compared with non-infected control; ++, p<0.01, +++p<0.001, planktonic compared with epithelial-associated CFU reduction (two-way ANOVA, Bonferroni post-test).
The CFU reduction caused by TV was not due to epithelial destruction as confirmed by light and electron microscopy (figure 1B). Cytopathic effects of TV were observed in older in vitro studies that used overly high parasite load (4–8×106 TV/mL).27 In our model, we applied a lower multiplicity of infection (∼1 TV cell per 5 epithelial cells or 4×105 TV/mL), which is much closer to the level of a typical infection diagnosed by wet-mount and thus more clinically relevant.28 At this dose, TV (the UR1 strain as well as many other virulent TV isolates) causes a robust inflammatory response within 24 h in the absence of cytotoxicity.15
To examine whether certain bacteria may have found refuge and remained viable within the epithelial cells, we performed experiments with antibiotics capable of killing extracellular bacteria using bacterial species that were less affected by TV: L crispatus, P bivia and A vaginae. Combined penicillin–streptomycin used routinely in cell culture completely suppressed the growth of these species while metronidazole suppressed P bivia only (figure 1C–E). The exposure of colonised epithelia to the penicillin/streptomycin reduced only the levels of L crispatus (figure 1F). P bivia and A vaginae survived poorly in the extracellular compartment (assessed by agar plating of cell culture supernatants) but maintained viability in epithelial cell lysates suggesting they were probably protected by being localised intracellularly (figure 1G,H).
To investigate whether alterations in bacterial colonisation was caused by TV or epithelial–TV interactions we conducted experiments in the presence or absence of TV comparing epithelia-associated with planktonic CFU (figure 1). TV significantly suppressed or destroyed lactobacilli and G vaginalis under planktonic conditions, and in the case of L acidophilus and L crispatus, these effects were enhanced in the presence of endocervical cells. P bivia was not significantly affected by TV under planktonic conditions and showed a tendency toward even higher numbers in colonised TV-infected epithelia. A vaginae did not grow under planktonic conditions but showed high numbers in the presence of colonised and TV-infected cells. Electron microscopy confirmed lack of cytotoxicity in our vaginal colonisation model, as previously shown,16 ,17 and also confirmed abundance of intracellular A vaginae (figure 1B) and P bivia (not shown). The ability of vaginal bacteria to reside inside the cervicovaginal epithelial cells is a phenomenon that warrants further investigation.
To investigate the effects of TV and its virulence factors on mucosal innate immunity in the presence of vaginal bacteria, we first measured levels of the pro-inflammatory chemokine IL-8, which is induced by TV, LPG, CPI-GC and TVV in the absence of bacteria;12 ,13 ,15 RANTES, which bridges innate to adaptive immunity via T cell recruitment and is secreted in response to viral infections and TVV-infected TV;15 and the anti-inflammatory microbicide SLPI, naturally abundant in human cervicovaginal secretions.28 For these analyses, we compared the three BV-associated spp. with L acidophilus and L crispatus, previously shown to lack pro-inflammatory activities.16 ,17
IL-8 (figure 2A) was universally upregulated (>twofold over medium control) by live TV, LPG, CPI-GC and TVV in the absence of bacteria; by each BV spp. or MALP-2; and only slightly by L crispatus and not by L acidophilus. The combination of TV with each BV spp. or MALP-2 significantly amplified the IL-8 response over TV alone (>twofold) or bacteria alone (especially A vaginae, >twofold) and the same was true when LPG or CPI-GC were applied. The increased IL-8 response was even more dramatic when TVV was combined with each BV spp. or MALP-2, suggesting a synergistic relationship: >20-fold over medium control, >fourfold over TVV alone, three- to fourfold over TV in the absence of bacteria or presence of lactobacilli, and >eightfold over lactobacilli alone.

Effect of Trichomonas vaginalis (TV) and bacteria on chemokine levels in an in vitro cervicovaginal epithelial model. (A–C) Levels of IL-8 (A), RANTES (B) and secretory leucocyte protease inhibitor (C) measured in epithelial-cell supernatants collected 48 h after bacterial colonisation as indicated (1–7, bacteria identified below panel C) and 24 h after exposure to TV isolate UR1 (TV), lipophosphoglycan (LPG), ceramide-phosphoinositol-glycan core (CPI-GC), Trichomonas vaginalis virus (TVV) virions or MALP-2. Data are means±SEM from duplicate cultures in one of five independent experiments. ***p<0.001, **p<0.01, *p<0.05, TV, LPG, CPI-GC and TVV different from medium control within each group, two-way ANOVA, Bonferroni post-test; +++p<0.001, ++ p<0.01, +p<0.05, medium+bacteria or medium+MALP-2 different from ‘no bacteria’ medium control, one-way ANOVA, Dunnett post-test. (D–G) Levels of IL-8 (D), RANTES (E), MIP3a (F) and IP-10 (G) assessed simultaneously by Meso Scale Discovery multiplex in epithelial-cell supernatants collected 48 h after colonisation with Lacidophilus acidophilus and Prevotella bivia and 24 h after exposure to TV (1–6, conditions identified below panel G). ***p<0.001, **p<0.01, different from medium control, +++p<0.001, ++p<0.01, P bivia different from P bivia+ TV, one-way ANOVA, Bonferroni post-test.
In the absence of bacteria, RANTES (figure 2B) was induced by TV and TVV but not LPG or CPI-GC, as expected from our previous work, in which RANTES upregulation was dependent on TVV dsRNA sensing in the context of multiple TV strains.15 RANTES was upregulated >fourfold by A vaginae or G vaginalis, less by P bivia (∼2-fold) and not by lactobacilli. A vaginae, G vaginalis or MALP-2 significantly amplified the RANTES response to TV (∼2-fold) and even more to TVV: 30- to 40-fold over medium control; five- to eightfold over TVV alone, TV in the absence of bacteria or TV in the presence of lactobacilli; and >18-fold over lactobacilli alone. In contrast to A vaginae and G vaginalis, P bivia suppressed TV- and TVV-induced RANTES. Purified LPG or CPI-GC suppressed the RANTES response to A vaginae or MALP-2.
SLPI (figure 2C) was significantly suppressed by live TV and even more by purified LPG or CPI-GC, both in sterile and bacteria-colonised epithelia. SLPI was slightly upregulated by G vaginalis, MALP-2 or TVV, but was nevertheless suppressed below baseline by TV, LPG or CPI-GC, even in the presence of G vaginalis or MALP-2.
To further investigate the dual (pro-inflammatory and immunosuppressive) effects of P bivia, we performed additional experiments in which TV infection was conducted in cervical cells colonised with either P bivia or L acidophilus, which had invariably shown a non-inflammatory profile (figure 2A–C and previous investigations16 17). Four chemokines were measured simultaneously by a Meso Scale Discovery multiplex. Again, similar to the results in figure 2A,B, P bivia induced vigorous IL-8 production and amplified IL-8 production induced by TV (figure 2D) but caused only a low RANTES response and suppressed TV-upregulated RANTES (figure 2E). MIP-3α was upregulated by either P bivia or TV alone in the presence or absence of bacteria, but together they yielded lower MIP-3α induction than P bivia alone (figure 2F). Similarly, IP-10 was upregulated by either P bivia or TV alone, but was completely suppressed by their combination (figure 2G). L acidophilus, in contrast, neither induced chemokines nor altered responses to TV (figure 2D–G).
Discussion
Our findings indicate that TV infection significantly reduced epithelia-associated Lactobacillus spp. but not BV spp. The results thus provide evidence for a cause–effect relationship between TV and BV and support clinical observations that trichomoniasis is associated with a specific vaginal bacterial community in reproductive-age women, characterised by a lack of significant numbers of lactobacilli and a higher proportion of strictly anaerobic organisms such as A vaginae, P bivia, G vaginalis and other BV spp.10
Since we frequently observed A vaginae and P bivia within epithelial cells, we hypothesise that some types of vaginal bacteria, particularly BV spp. and especially A vaginae, may escape the hostile vaginal environment and attacks by TV by localising inside epithelial cells. In our experiments, antibiotics at doses that killed planktonic or extracellular bacteria had less effect on P bivia and did not reduce A vaginae CFU obtained from epithelial lysates, suggesting these bacteria may remain viable at significant numbers within epithelial cells. It cannot be excluded, though, that these and other bacteria might form a surface biofilm that makes them more resistant to antibiotic treatment and perhaps also to TV or TV products. Our findings thus warrant further investigations that may suggest novel potential mechanisms for managing recurrent and antibiotic-resistant BV.
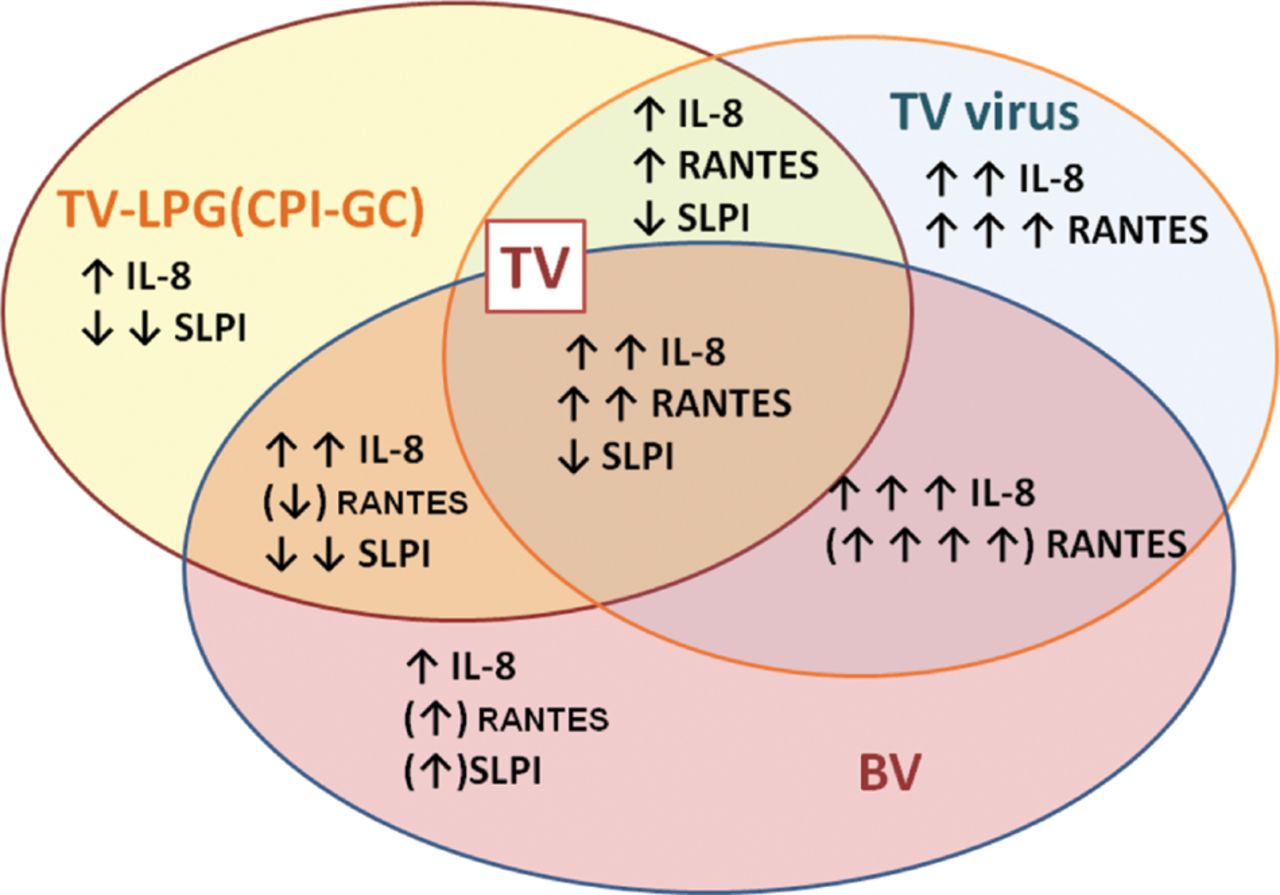
The combination of BV spp. with TV significantly affected the host immune response to the individual TV virulence factors LPG and TVV as well as to the live parasite as a whole (schematised in figure 3). Importantly, our experimental results show also a remarkable causative relationship between altered epithelial responses and the type of vaginal bacteria populating the epithelia at the time of TV infection, with Lactobacillus spp. lacking and BV spp. possessing the ability to alter different aspects of LPG- or TVV-driven responses.
{kind=link}
{kind=link}
{kind=link}
Schematic presentation of chemokine response and suppressed secretory leucocyte protease inhibitor in response to Trichomonas vaginalis (TV) virulence factors and combined bacterial vaginosis (BV)-TV infection. Arrows show magnitude and direction of change. Arrows in parentheses indicate responses dependent on type of BV bacteria.
The fact that TV teams up with A vaginae and G vaginalis to induce much higher chemokine responses (represented here by IL-8 and RANTES) on the background of significantly reduced SLPI due to LPG/CPI-GC signalling suggests a mechanism for inflammatory damage accompanied by recruitment of CD4 cells and weakened antiviral barrier thus facilitating viral co-infections including herpes simplex virus, human papillomavirus and HIV. SLPI is an innate-immune mediator with direct virucidal effects, which is produced and stored at abundant levels in mucosal epithelial cells and keratinocytes.16 ,17 SLPI is also capable of dampening inflammatory responses to LPS.29 We have recently shown that vaginal SLPI levels decrease in women with trichomoniasis in a manner dependent on parasite load.28 Higher vaginal SLPI levels may have protective anti-HIV activity as suggested by studies in HIV controllers.30
The immunosuppressive effects of P bivia warrant further investigation. Suppression of IP-10, MIP-3α and RANTES may serve to evade the arm linking innate to adaptive immunity since those chemokines in combination are particularly important for recruitment of antigen-presenting dendritic and T cells to the site of microbial invasion. Our data also suggest that the effects of TV on the vaginal immunobiome depend on which BV spp. are prevalent at the time of TV inoculation. The fact that TVV causes a tremendous (in some cases over 30-fold) amplification of the inflammatory reaction to TV is a point of particular concern and suggests the need to include antiviral and anti-inflammatory components in the combined therapeutic–preventative approach to trichomoniasis leading to or occurring in concert with BV.
Conclusions
Our findings have important implications for understanding how TV and its major virulence factors LPG and TVV affect the mucosal immuno/microbiome and for designing better therapeutic approaches for combining antiparasitic therapy with attempts to restore the normal microbiome by introducing Lactobacillus-based probiotics and preventing TV-attributable BV and inflammatory sequelae. Further experimental and clinical studies should follow for identifying the molecular targets for novel drug therapies preventing additive or synergistic pro-inflammatory and immunosuppressive effects of TV, TVV and BV in concert.
Key messages
-
Trichomonas vaginalis reduced epithelia-associated Lactobacillus spp. but not bacterial vaginosis (BV) spp.
-
T vaginalis and BV species in concert amplified pro-inflammatory and suppressed protective innate-immune responses.
-
Virulence factors of T vaginalis that altered epithelial responses in concert with BV species include the surface lypophosphoglycan and endosymbiotic dsRNA viruses.
-
Future therapeutic/preventative approaches should combine antiparasitic therapy with microbiome restoration and target the pro-inflammatory and immunosuppressive effects of T vaginalis and BV.
Acknowledgments
The authors thank Maria Ericsson at the Harvard Medical School Electron Microscopy Facility for technical assistance.
References
Footnotes
ORB, HSY and TF all contributed equally.
-
Handling editor David Lewis.
-
Contributors RNF conceived and directed the co-infection model and concept development, cloned the UR1 isolate, wrote the manuscript, and coordinated reviews and coauthors’ contributions. ABO, BNS and MLN contributed to concept development and provided critical review of results and interpretations, ABO provided microbiology expertise and characterised vaginal bacteria isolates; BNS provided the original UR1 isolate and expertise in Trichomonas vaginalis biochemistry; and MLN provided virology expertise and TVV virions. ORB, HSY and TF conducted the experiments, cell viability assessment and data analyses; MD characterised and expanded bacterial isolates; HYD, BF, OB and TF performed bacterial cultures; ORB and HYD performed immunoassays; DHB and TF cultured T vaginalis, and GRH and BNS purified LPG and CPI-GC; and YT purified the TVV1 virions. All authors read, provided critical comments and agreed with the final manuscript content.
-
Funding This work was supported by the National Institute of Allergy and Infectious Diseases (1RC1AI086788-01, 1R56AI091889-01A1 and 5R01AI079085) and the National Institute of Child Health and Human Development (R21HD054451).
-
Competing interests None.
-
Ethics approval The original protocol for collecting Trichomonas vaginalis isolates with patients' informed consent was approved by the IRB boards at SUNY Upstate Medical University (Syracuse, New York, USA) and Brigham and Women's Hospital (Boston, Massachusetts, USA).
-
Provenance and peer review Not commissioned; externally peer reviewed.