Article Text

Abstract
Objectives In 2006 a new variant of Chlamydia trachomatis (nvCT), with a deletion in the cryptic plasmid, was reported in Sweden. This deletion included the targets for the genetic diagnostic systems used in many clinical laboratories and resulted in thousands of false-negative results. The aim of this study was to characterise consecutive Chlamydia tissue culture-positive samples from 2006 in Örebro County, after identification of the nvCT, and to compare the results from samples collected in the same county in 1999–2000. The study also aimed to evaluate the discriminatory capacity of multilocus sequence typing (MLST) compared with ompA sequencing.
Methods ompA sequencing and MLST was used to characterise 100 consecutive Chlamydia tissue culture-positive samples.
Results A significant (p<0.001) increase of genotype E, from 47% in 1999–2000 to 69% in 2006, was detected. All 41 nvCT isolates from 2006 displayed an identical ompA genotype E and MLST profile. Excluding the nvCT isolates, the distribution of ompA genotypes is similar to the genotyping results from 1999–2000. Among the wild-type genotype E isolates from 2006, 14 unique MLST sequence types were obtained from 26 isolates while they were identical in ompA genotyping. The discriminatory power (D) of C trachomatis strains in this material was 83.5% using the MLST system compared with 49.5% utilising ompA sequencing.
Conclusion In all, MLST enables improved studies of the molecular epidemiology of C trachomatis. All nvCT isolates from 2006 displayed an identical ompA genotype E and MLST profile, which strongly indicates a clonal spread of the nvCT.
- Chlamydia trachomatis
- genotype
- laboratory diagnosis
- molecular typing
- multilocus sequence typing
- mutant
- ompA
- MLST
- multilocus sequence typing
- NAAT
- nucleic acid amplification test
- nvCT
- new variant of Chlamydia trachomatis
- ompA
- outer membrane protein A
Statistics from Altmetric.com
- Chlamydia trachomatis
- genotype
- laboratory diagnosis
- molecular typing
- multilocus sequence typing
- mutant
- ompA
- MLST
- multilocus sequence typing
- NAAT
- nucleic acid amplification test
- nvCT
- new variant of Chlamydia trachomatis
- ompA
- outer membrane protein A
In 2006 it was reported in Sweden that a proportion of the sexually transmitted Chlamydia trachomatis infections could not be detected using standard nucleic acid amplification tests (NAAT) manufactured by Abbott Laboratories (Abbot Park, Illinois, USA) and Roche Diagnostics (Pleasanton, California, USA) due to a deletion in the cryptic plasmid.1 At that time these manufacturers supplied approximately 70% of all C trachomatis tests in Sweden,2 and the diagnostic systems Abbott m2000, Abbott and Cobas Amplicor/TaqMan48, Roche used targets in the deleted region of the cryptic plasmid. Almost all other C trachomatis tests in Sweden were performed with BD ProbeTec ET (Becton Dickinson, Franklin Lakes, New Jersey, USA), which uses a target sequence in another region of the same plasmid.3 To date, the new variant of C trachomatis (nvCT) has been reported in most Swedish counties, with published proportions ranging from approximately 10% to 65% of the true positive C trachomatis samples.4 5
Characterisation of C trachomatis ompA (omp1) genotypes has been widely used as a tool in epidemiological studies.6 7 However, in unselected or less selected populations it has been shown to provide limited discrimination,8–12 especially for the predominating genotype E that in most studies comprises over 40% of the C trachomatis cases.6 7 Therefore, more discriminative methods for examination of the molecular epidemiology of chlamydial infections are of crucial importance and both a multilocus sequence typing (MLST) system13 and a system based on tandem repeats combined with ompA sequencing14 have been developed.
In Örebro County, which comprises approximately 275 000 inhabitants, all C trachomatis samples are analysed at Örebro University Hospital. During the past decade, the rates of reported C trachomatis cases per 100 000 inhabitants in the county have been relatively similar to the national rates, which increased between 1997 and 2005. However, the rate per 100 000 inhabitants showed a decrease, from 336 in 2005 to 311 in 2006.5 The diagnostic methods used at that time were NAAT, Cobas Amplicor (Roche), and culture on McCoy cells. In 2005, the percentage of C trachomatis-positive samples was 5.4% using tissue culture and using NAAT 7.1%. The corresponding figures for 2006 were 5.6% and 6.5%, respectively. Consequently, the level of PCR positivity decreased slightly, whereas the positivity level using culture diagnostics was stable.
In a previous study in Örebro County in 1999–2000, genotyping of genital C trachomatis infections based on ompA (omp1) sequencing was performed in specimens collected at the Sexually Transmitted Diseases Clinic, Örebro University Hospital, during a one-year period.10
The aims of the present study were to ompA genotype 100 consecutive C trachomatis culture positive samples from 2006 including the nvCT, and compare the results with the genotyping results from the one-year study performed in the same county in 1999–2000, as well as to characterise the 2006 isolates with a new MLST method.13 The study also aimed to evaluate the discriminatory capacity of MLST compared with ompA sequencing.
Materials and methods
Clinical specimens
One hundred consecutive C trachomatis tissue culture-positive samples from routine diagnostics from 5 October to December 2006 were genotyped using ompA sequencing and MLST. These samples also included nvCT-positive samples, which were found using Cobas Amplicor (negative results) and mutant-specific real-time PCR.15 In addition, the results of the ompA sequencing of 237 C trachomatis-positive clinical specimens, collected during a one-year period in 1999–2000,10 were used for comparison, and a subset of 53 genotype E samples were also examined by MLST.
The age of the individuals examined was recorded from referral sheets to the laboratory.
ompA typing
Sequence determination was according to our previously described method.10
Multilocus sequence typing
The system comprises five target regions (figure 1) and was performed as previously described13 with a minor modification. This consisted of the pbpB gene being amplified as two separate fragments, using the primer pairs pbpB1F/pbpB823R (5′–TATATGAAAAGAAAACGACGCACC–3′ and 5′–CAGCATAGATCGCTTGCCTAT–3′) and pbpB1455F/pbpB2366R (5′–GGTCTCGTTTTTGATGTTCTATTC–3′ and 5′–TGGTCAGAAAGATGCTGCACA–3′), respectively. The pbpB gene was amplified using the same cycling parameters as for the other regions.

Schematic overview of the five multilocus sequence typing (MLST) regions. Each region is named after the target gene. The hctB has repetitive elements (grey) and the pbpB was analysed in two reactions (pbpB1F/pbpB823R and pbpB1455F/pbpB2366R).
Discriminatory power
The discriminatory power (D) was determined as previously described by Hunter and Gaston,16 for the ompA sequencing, the MLST system and the combination of both methods. D is the index of the discriminatory power expressed in a percentage. As D must be based on epidemiologically unrelated isolates, only 97 isolates from 2006 were used in this calculation, because three isolates were from sexual contacts.
Statistics
To test the difference of proportion with genotype E in 1999–2000 and 2006 we used the Pearson χ2 test. The significance level was set at 5% (two-sided). Statistical analyses were performed using SPSS software V.15.0.
Results
ompA genotyping
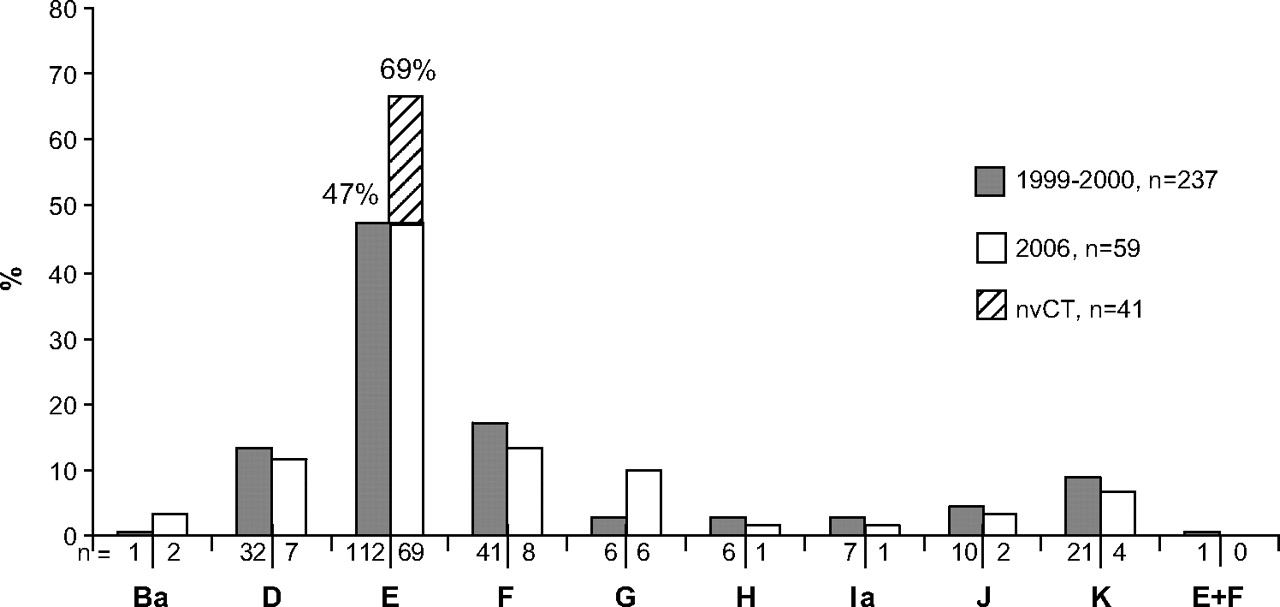
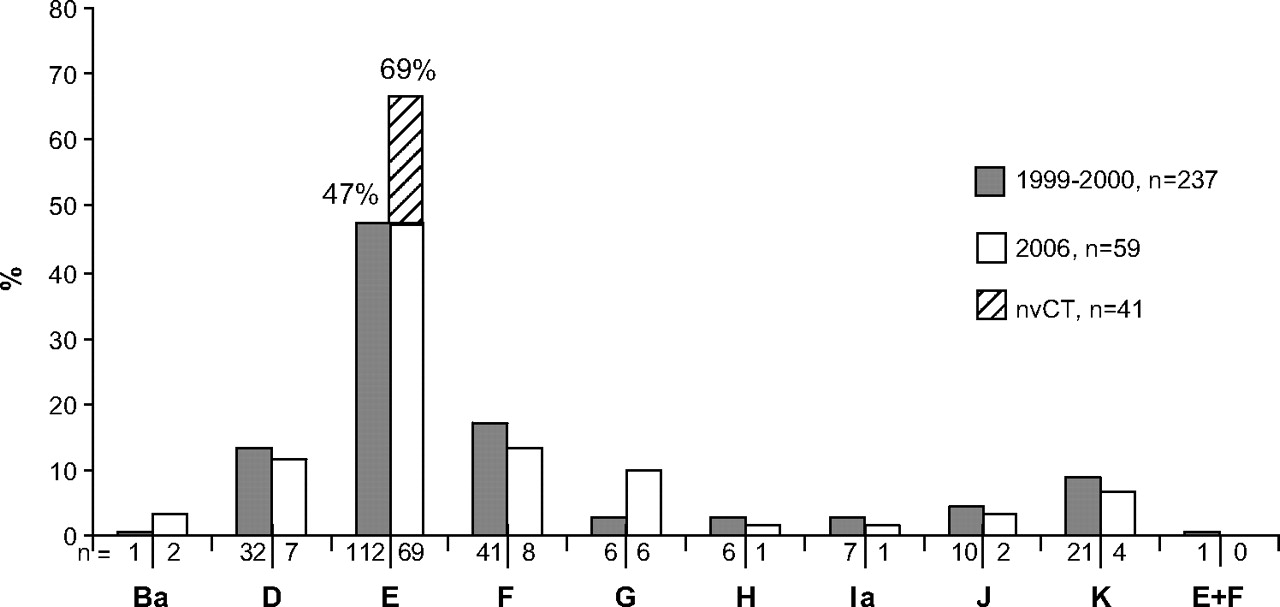
Among the 100 consecutive Chlamydia tissue culture-positive samples in the 2006 material, there were 69 (69.0%) genotype E isolates, with an identical ompA sequence. Genotype E was thus significantly more prevalent (p<0.001) than in 1999–2000 (47.3%, figure 2).
{kind=link}
{kind=link}
ompA genotype distribution of 237 urogenital Chlamydia trachomatis isolates from a one-year study in Örebro, Sweden, in 1999–200010 compared with the distribution of genotypes in consecutive urogenital isolates (n=100) in the same county from October to December 2006. All 41 new variant (nvCT) cases were excluded before comparison, but are denoted in the figure as genotype E.
Among the genotype E isolates cultured in 2006, all 41 of the nvCT displayed identical ompA sequence compared with the reference strain E/Bour (GenBank no X52557). However, three of these nvCT-positive samples were also Cobas Amplicor positive, indicating multiple infections also including wild-type C trachomatis (wtCT). Excluding the 41 nvCT isolates, the distribution of ompA genotypes (n=59) was as follows: genotype B (3.4%), D (11.9%), E (47.5%), F (13.6%), G (10.2%), H (1.7%), Ia (1.7%), J (3.4%) and K (6.8%), which is similar to the genotyping results from 1999–2000 (see figure 2). The mean age in 2006 was lower (21.1 years, range 15.8–35.4, median 20.1 years) for the 41 patients with nvCT compared with the 59 patients with wtCT (24.3 years, range 15.4–56.3, median 21.4 years). In the one-year period in 1999–2000 the mean age was 27.2 years (range 13.5–63.5).
Multilocus sequence typing
The distribution of the MLST profiles from 2006 is shown in tables 1 and 2. Briefly, complete MLST profiles were obtained from 26 out of 28 wtCT genotype E isolates, and among these 14 (58%) unique sequence types were identified. In contrast, all nvCT isolates displayed an identical and unique MLST profile: 21 (target region hctB), 19 (CT058), 1 (CT144), 2 (CT172) and 1 (pbpB). However, one isolate had a too short sequence product and an inconclusive result in pbpB, but was typeable to be 1, 2 or 38.
Discrimination of 69 genotype E wtCT and nvCT isolates from 2006 using MLST and ompA sequencing
Distribution of MLST profiles and C trachomatis ompA genotypes in 31 clinical isolates from 2006, excluding the genotype E isolates described in table 1
The MLST profiles of the 31 isolates that displayed other ompA genotypes than genotype E are shown in table 2. In short, among the genotype D isolates (n=7), the MLST method and the ompA genotyping both identified three different genetic variants. For the genotype F isolates (n=8) the ompA sequences were identical, but the MLST method showed four different sequence types and one additional isolate was not typeable in hctB. All genotype G isolates (n=6) showed different sequence types, while two ompA sequences were identified. Two genotype J isolates were identified, displaying identical ompA sequences but different sequence types. All genotype K isolates (n=4) displayed an identical ompA sequence, but three different sequence types. It is notable that in four of the ompA genotype D and three of the ompA genotype F isolates the MLST profile was identical (5, 19, 7, 1, and 4).
Out of 53 genotype E isolates from the 1999–2000 material, with an identical ompA sequence, we only managed to identify four complete MLST profiles (data not shown). However, we were able to type the target region hctB in 16 of the 53 isolates, but none displayed the allele variant number 21, which was represented in all nvCT in 2006 (see table 1).
Discriminatory power
The D was calculated using 97 unrelated isolates. D for ompA sequencing in this material was 49.5% and for the MLST method it was 83.5%. With a combination of both methods, D=83.7%.
Discussion
Our genotyping study of 100 consecutive C trachomatis isolates shows that the outbreak of nvCT is of clonal nature and that MLST provides a substantially higher discriminatory power than ompA sequencing in genotype E isolates, resulting in more than a threefold higher number of variants compared with ompA sequencing when nvCT cases are excluded.
We have previously shown that, in principle, our MLST system provides high resolution for genotyping of selected C trachomatis cases.13 In the present study MLST was further challenged by typing 100 consecutive samples and compared with ompA genotyping. This investigation confirmed that MLST is also superior to ompA sequencing in discrimination capacity for unselected specimen collections.
For genotypes E, F, G and K, comprising 87% of all C trachomatis cases, the discriminatory capacity of MLST was substantially higher than when utilising ompA sequencing. For the seven genotype D isolates both typing systems resulted in three genetic variants, which is in contrast to a previous study in which genotype D samples showed five different ompA sequences, but 11 divergent MLST sequence types.17 However, this was in a study of men who have sex with men and the high number of partners they had may explain the different outcome. For the other genotypes (B, H, I and J) no difference between the systems could be seen, but the number of cases was also too low to enable a valid comparison.
It is also noteworthy that using the five MLST targets and ompA sequencing in combination was beneficial, as four of the ompA genotype D and three of the ompA genotype F isolates displayed a single identical sequence type. This would also be useful for comparisons with many previous and presumably future studies using ompA genotyping. Nevertheless, overall ompA sequencing only slightly increased the number of genetic variants identified.
We found that all nvCT isolates were of genotype E, with an identical ompA sequence, and displayed an identical and unique MLST profile, which strongly indicates a clonal spread of the new variant. In contrast, among 26 of 28 wtCT genotype E isolates, 14 (58%) unique MLST types were obtained. The mean age of patients infected with nvCT was lower than that of patients harbouring wild-type strains. This was not surprising, considering that young persons in Sweden are more sexually active and have more partners, and thus nvCT is more rapidly spread in younger age groups. The nvCT has been detected in high proportions all over Sweden,18–20 but surprisingly few cases have been found in neighbouring countries or other countries.2 21–23 Furthermore, in a recent study the MLST analysis of 48 arbitrarily selected nvCT isolates from four countries had one single sequence type, which indicates a clonal spread.2 The present investigation of a local study population confirmed these findings. It also indicates that although the MLST system has high resolution, there is no obvious risk that the system is too variable and that it would give misleading results when used for molecular epidemiology of sexual networks.
At present it is not known when or where this nvCT appeared and started to be transmitted in Sweden. However, in our study in 1999–2000,10 one suspected mutant (of 237 positive samples derived from a total of 2195 patients), which was positive in conventional ompA gene PCR but repeatedly negative in Cobas Amplicor, was identified. This specimen was also determined as genotype E, that is, as the present nvCT, but 47% of the samples in that previous study were genotype E. At that time, this Cobas Amplicor-negative isolate was considered a plasmid-free C trachomatis strain.24
Efforts have been made to apply the present MLST method to C trachomatis specimens from the previous study in 1999–2000.10 However, complete MLST profiles were obtained in only a limited number of cases. This is explained by the storage of the samples at –20°C and previous repeated freeze thawing, resulting in DNA degradation.18 25 Nevertheless, we managed to show that in one target region (hctB) the allele variant for genotype E in 1999–2000 differed when compared with that in 2006. It is not surprising that a fluctuation of specific C trachomatis strains predominates in a population, but as conventional ompA typing has lower differentiating capacity this is not easily seen.
There are limitations in our study. First, we apparently only succeeded in analysing a few samples from 1999–2000 in the MLST system. This reduces the possibilities for comparing variation of sequence types over time. In addition, the number of isolates analysed in 2006 (n=100) also means that statistical changes are difficult to obtain. Second, clinical and epidemiological data for the Chlamydia-infected individuals in the study were mainly lacking. Analysis of the spread of specific C trachomatis strains in sexual networks was thus not possible and was also beyond the aims of the study.
Although for two decades genotyping of C trachomatis has been restricted almost entirely to ompA sequencing, new methods with other targets have recently been reported. One system combines ompA sequencing with analysis of tandem repeats.14 An MLST system intended for tracking evolutionary changes has also been published.26 The advent of new multitarget typing systems provides more information and highlights the need for standardised nomenclature systems.
In conclusion, the identical MLST profile of all nvCT isolates strongly indicates a clonal spread in the population, as shown by this study, in which a significant (p<0.001) increase in genotype E from 47% in 1999–2000 to 69% in 2006 was detected. Excluding the nvCT isolates, the distribution of ompA genotypes is similar to the genotyping results from 1999–2000. The overall discrimination of C trachomatis isolates was high using the present MLST system. Accordingly, the MLST system enables improved molecular epidemiological characterisation of sexually transmitted Chlamydia infections.
Key messages
The MLST method provides a substantially greater discrimination between C trachomatis isolates than ompA sequencing.
All nvCT isolates displayed an identical ompA genotype E and MLST profile, which strongly indicates a clonal spread of nvCT.
When excluding the nvCT isolates, the distribution of C trachomatis ompA genotypes is similar to the genotyping results from the same county in Sweden in 1999–2000.
Acknowledgments
The authors would like to thank Anders Bäckman, Thomas Morin and Sara Ödmark for technical assistance with the genetic analyses.
References
Footnotes
The work of this manuscript is part of the goals described in the European Framework Programme 6 (FP6) funded EpiGenChlamydia Consortium (EU FP6 LSHG-CT-2007-037637) a co-ordination actions in functional genomics research entitled: ‘Contribution of molecular epidemiology and host–pathogen genomics to understand Chlamydia trachomatis disease’ (see additional information at http://www.EpiGenChlamydia.EU).
Funding This work was supported by grants from the Örebro Medical Research Foundation, Örebro University Hospital, the Uppsala-Örebro Regional Research Council, the National Board of Health and Welfare, Stockholm, and the National Institute for Public Health, Östersund, Sweden.
Competing interests None.
Provenance and peer review Not commissioned; externally peer reviewed.
Linked Articles
- Whistlestop tour
- Correction